Innervation is required for sense organ development in the lateral line system of adult fish
Wada H, Dambly-Chaudière C, Kawakami K, Ghysen A. (2013)
Proc. Natl. Acad. Sci. USA 110, 5659-5664.
Innervation is required for sense organ development in the lateral line system of adult fish
Wada H, Dambly-Chaudière C, Kawakami K, Ghysen A. (2013)
Proc. Natl. Acad. Sci. USA 110, 5659-5664.
軸索投射が、感覚器の細胞増殖に必要
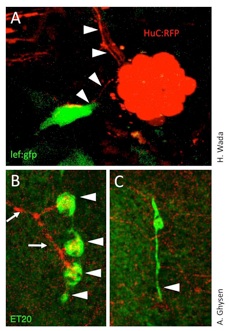
感覚器には必ず感覚神経軸索が分布し、シグナルを脳に伝えています。感覚器からはさまざまな因子が分泌され、神経軸索を誘引・増殖することが知られていますが、軸索の役割についてはよくわかっていません。我々は、魚の側線器官(感丘)の細胞増殖は、軸索によって制御されることを示しました。
増殖する感丘では、Wntシグナル活性が細胞増殖を制御していると考えられます(トピック1)。Wnt活性が高い増殖細胞は必ず、分岐した軸索に投射されます(図A,矢頭)。神経細胞または軸索をレーザー照射装置により除去すると、感丘は出芽するにもかかわらず、細胞増殖が起きないことを示しました(図B)。つまり、軸索はWnt活性を亢進することによって感丘の細胞増殖を制御しています。軸索と感覚器の密接な相互作用を示しています。
(モンペリエ大学、アラン・ギーセン博士との共同研究)
Dermal morphogenesis controls lateral line patterning during postembryonic development of teleost fish
Wada H, Ghysen A, Satou C, Higashijima S, Kawakami K, Hamaguchi S, Sakaizumi M. (2010)
Dev. Biol. 340, 583-594.
側線神経系が多様なパターンを生み出す仕組み
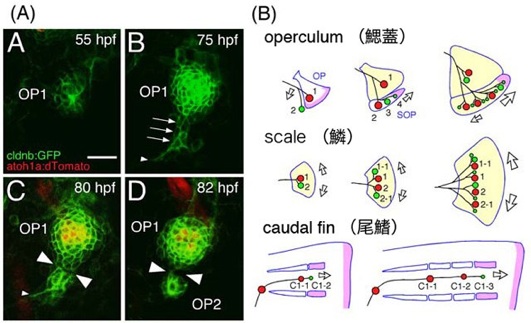
神経系は、脳の領域や動物の種類によって、著しく異なったパターンを示します。私たちは、魚が水の流れを感じる側線神経系に着目し、神経回路が多様なパターンを形成する仕組みを調べました。側線神経は、体表面に分布する感覚受容体(感丘)を支配します。魚の成長過程で、感丘の一部の細胞が遊走し、分裂し、さらに凝集することによって、新しい感丘を形成します(図A)。次に、感丘は定期的に「出芽」を繰り返すことによって数を増し、側線神経軸索は枝分かれをして、新しく生まれた感丘を追って伸びることを示しました。また、鰓蓋の形成に異常を示す魚を調べた結果、感丘の位置(すなわち側線パターン)は、鰓蓋骨の位置によって決まることを示しました。このような表皮の下に形成される骨(皮骨)と側線パターンの相関は、鱗や尾鰭でも見られました(図B)。さらに、魚種による側線パターンの違いのいくつかは、その魚種に特徴的な皮骨形成によって説明できることを示しました。以上の結果は、側線パターンが、皮骨形成によって制御されていることを示すとともに、側線神経系が多様なパターンを生み出す仕組みの一端を示しています。
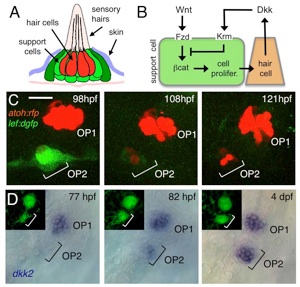
Wnt/Dkk negative feedback regulates sensory organ size in zebrafish
Wada H, Ghysen A, Asakawa K, Abe G, Ishitani T, Kawakami K (2013) Curr. Biol. 23: 1559-1565
負のフィードバックにより感覚器官のサイズが制御される
体の様々な器官は幹細胞を持ち、self-renewalによって細胞が新しく置き換わったり、受けた損傷を修復したりして器官を一定の大きさに保ちます。どのようにして、器官は一定のサイズを認識するのでしょうか。
ゼブラフィッシュの側線器官(感丘)は、中央の感覚細胞(有毛細胞)とそれを取り囲む支持細胞からなります(図A)。感丘は、出芽によって増え、クラスターを形成しますが、それぞれの感丘は、同じ大きさをしています。我々は、Wntシグナル活性が、増殖細胞で高いこと(図C)、感覚細胞がWntシグナルの抑制因子Dkk2を発現することを見出しました(図D)。Wntシグナル、Dkkシグナル、それぞれの機能阻害・獲得実験から、Wntシグナルは細胞の増殖を促進し、DkkはWntシグナルを抑制することによって細胞増殖を阻害することがわかりました(図B)。つまり、Wnt/Dkkは負のフィードバック・ループを形成し、器官の大きさを一定に保つことが示されました。負のフィードバックは、多くの恒常的システムに必須のコンポーネントです。本研究は、器官サイズ調節における、負のフィードバックの分子機構を明らかにしました。
この論文は「Faculty of 1000」に選ばれました。
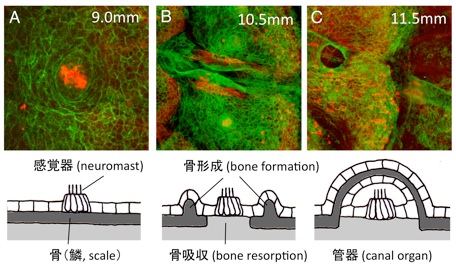

動物の皮膚はさまざまな組織からなる複雑な構造をしています。皮膚の感覚器は、決まった位置に決まったパターンで存在することによって、適切な感覚受容を行っています。これらの感覚器は、どのように形成されるのでしょうか。
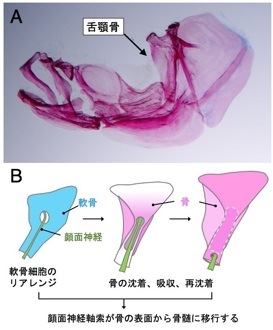
ゼブラフィッシュの側線器官(感丘)は、稚魚期には体表にありますが(図A)、魚の成長過程において、骨組織であるウロコの中に取り込まれます(管器、図C)。我々は、この移行過程において、骨形成と骨吸収(骨リモデリング)が行われていることを明らかにしました(図B)。さらに、ウロコの移植実験と突然変異体解析から、(1)側線器官が骨リモデリングに必要であること、また、(2)骨リモデリングは側線器官の成長に必要であること、を示しました。これらの結果は、感覚器(神経組織)と骨(結合組織)は、密接に相互作用しながら、形態形成を行っていることを示しています。
骨組織の恒常性の維持は、生体にとって重要な役割を持っています。骨形成と骨吸収のバランスが崩れることによって、骨粗鬆症などの骨疾患が生じます。魚のウロコは、体表面に露出した骨組織であるため、その形成過程を生きたまま観察することができます。骨リモデリング過程における組織間の相互作用を明らかにすることが今後の課題です。
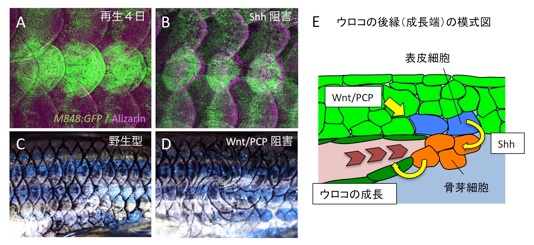

魚のウロコは平たい骨でできていて、薄い表皮の下に決まったパターンで並んでいます。ウロコは脱落してもわずか4日後には生え替わります(図A)。この高い再生能力の仕組みを調べることによって、骨の形づくりや再生の研究に役立てることができます。
この研究では、まず、ウロコの成長末端にある骨芽細胞(骨を作る細胞)が、Shh遺伝子を発現する表皮細胞と接していることを示しました(図E)。次に、Shh遺伝子を薬剤によって阻害すると、骨芽細胞の形成が抑制され、ウロコの再生が阻害されました(図B)。さらに、Shh遺伝子の発現が、表皮細胞の平面極性(細胞が一定方向に並ぶ現象)によって制御されることを示しました。表皮細胞の平面極性をつかさどるWnt/PCPシグナルを阻害すると、Shh遺伝子の発現パターンが変わり、ウロコの成長する方向も変わりました(図D)。
動物の体を支えるためには、骨の形や成長(パターン)が正確にコントロールされる必要があります。この研究によって、骨のパターンを制御する分子メカニズムの一端が明らかになりました。
↓ 日本骨代謝学会で紹介

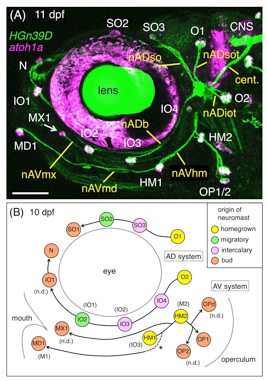
魚の頭部の側線系は、魚種によって大きく異なります。これは、それぞれの魚が異なる環境に生息し、周囲の水の流れを感知しているからです。頭部の側線パターンは非常に複雑であるため、どのように発生するのかよく分かっていませんでした。この研究では、ゼブラフィッシュを用いて、側線器官(ニューロマスト・感丘)、神経軸索、感覚細胞、軟骨細胞を蛍光タンパク質(GFP、RFP)で標識することで、それらの発生段階を詳しく記載しました(図A)。
その結果、頭部に分布するニューロマストは、4つの異なる仕組みで生じることが分かりました(図B)。1.側線原基が生まれた場所で形成されるもの(黄色)、2.側線原基が移動して生じるもの(緑)、3.幹細胞が増殖して生じるもの(ピンク)、4.側線原基が「芽」を出して生じるもの(オレンジ)。さらに、顎の決まった場所にニューロマストが生じるためには、舌弓(鰓や顎の構造の1つ)に隣接する細胞(神経堤細胞)からの分泌因子Rspo2が必要であることを見出しました。
この論文は、進化の過程で、魚がどのように異なる側線パターンを生み出し、環境に適応してきたのかを理解する上で、基盤となる研究です。